How a Hidden Proton Channel Sabotages Brain Cells in Parkinson’s
TL;DR: A newly decoded proton channel called TMEM175 lies dormant in lysosomes until acid arrives—then it opens wide and floods the cell with hydrogen ions, disrupting the delicate pH balance linked to Parkinson’s disease and other neurodegeneration.
Your cells run a 24/7 recycling system inside tiny acid baths called lysosomes — and when those baths lose their pH balance, the brain pays the price. A new study reveals that the ion channel TMEM175, long linked to Parkinson’s disease risk, has a hidden proton-gating mechanism that could be the key to understanding why.
Key Findings
- Acidification triggers explosive H+ influx: When pH dropped from neutral (7.4) to acidic (4.7) on the luminal side of TMEM175, inward and outward currents increased continuously, indicating the channel rapidly admitted hydrogen ions.
- Protons collapse the pH gradient in minutes: The H+ current was so intense that it quickly dissipated the lysosome’s acid gradient—the very condition it needed to function—a process that continued for 90 seconds or longer.
- Reversal potential drops ~100 mV below theoretical H+ equilibrium: Even without competing cations, the measured reversal potential lagged far behind what pure H+ selectivity would predict, indicating rapid local pH changes around the pore.
- H57 is the critical gatekeeper residue: A single histidine at position 57 (H57) sits on the inside of the channel pore and forms salt bridges that modulate both gating and ion selectivity.
- H57Y mutation cuts H+ flow by fourfold: Mutating H57 to tyrosine (H57Y) reduced proton permeability by 75% and knocked K+ permeability down by a factor of 15, proving H57’s central role.
- H57Y lysosomes show blunted acid-responsiveness: Whole-lysosome recordings confirmed that lysosomes carrying the H57Y mutation had only slightly elevated currents compared to controls, even during acidification.
Source: PNAS (2026) | Schulze et al.
The pH Collapse Problem
The research team, led by Tobias Schulze and colleagues at multiple European institutions, used patch-clamp electrophysiology—a gold-standard technique for measuring single-channel currents—to directly observe what TMEM175 does when lysosomes turn acidic. The finding was stark: the channel doesn’t just passively let protons through; it actively pumps them in, and does so with such ferocity that it erodes the very pH gradient the lysosome needs to survive.
This is a functional paradox. The lysosome must be acid to activate TMEM175. But once activated, TMEM175 shuts down the acidity that activated it. The researchers quantified this: in less than 90 seconds, the reversal potential (a measure of the transmembrane ion gradient) drifted toward zero, indicating the proton concentration difference had nearly vanished.
Finding the Molecular Culprit
To understand how TMEM175 achieved this feat, the team turned to molecular dynamics simulations—computational models that show how amino acids move and interact inside the channel pore. They tested which residues might control the pH-dependence and ion selectivity of TMEM175.
Histidine at position 57 (H57) emerged as the key. This amino acid sits on the luminal-facing side of the open channel, and its chemical behavior changes with pH. At neutral pH, H57 is uncharged; at acidic pH, it gains a positive charge. The simulations showed that protonated H57 forms salt bridges—electrostatic attractions—with nearby aspartate (D279) and glutamate (E282) residues, stabilizing the channel in a state that favors proton influx.
To test this prediction, the researchers created a mutant: H57Y (replacing histidine with tyrosine). Tyrosine cannot change its charge with pH, so it should break the pH-sensing mechanism.
The Mutation Proves the Point
The H57Y mutant delivered compelling evidence. In whole-cell recordings, H57Y cut proton permeability by a factor of four compared to wild-type TMEM175. Equally striking: potassium permeability plummeted by 15-fold. This wasn’t a minor tweak—it was a near-complete loss of function.
The researchers then moved to whole-lysosome patch-clamp recordings, a more physiologically relevant setup. Lysosomes carrying the H57Y mutation showed only slightly higher currents even during acidification, far below the sustained increases seen in wild-type lysosomes. The pH-sensing behavior was gone.
Why This Matters for Parkinson’s Disease
TMEM175 dysfunction is linked to Parkinson’s disease. Genetic variants in TMEM175 show up in large genome-wide association studies (GWAS) as risk factors for the disease. The mechanism is still being pieced together, but the broad logic is clear: if TMEM175 fails to regulate lysosomal pH properly, waste products and damaged proteins accumulate. In neurons, this junk piles up until the cell dies.
Now researchers understand one side of that story. A normal TMEM175 channel can be too efficient—it admits so many protons that it collapses the pH gradient and sabotages lysosomal function. A mutant TMEM175, especially one affecting H57, loses this destructive capacity but also loses protective capacity. The problem is balance.
The finding also clarifies what makes TMEM175 unusual. Most proton channels attempt to maintain electrochemical gradients. TMEM175 appears designed to collapse them—perhaps as a pressure-relief valve when lysosomes become too acidic, or to enable pH-dependent recycling pathways. But if this collapse mechanism misfires, the lysosome becomes a ghost organelle: unable to digest, unable to maintain itself, and unable to protect the cell from neurotoxic aggregates.
A Cautionary Tale in Channel Design
This study reveals something deeper about how ion channels evolved. TMEM175 is elegant: a single amino acid (H57) acts as both a pH-sensor and a gating control. When acid arrives, H57 gains a proton charge, locks into place via salt bridges, and opens the floodgates. But elegance carries risk. A tiny mutation, or a genetic variant that slightly shifts H57’s pKa (the pH at which it changes charge), could turn a protective valve into a broken dam.
For the millions of people at risk for Parkinson’s disease, these discoveries hint at new therapeutic angles. Instead of trying to block TMEM175 outright—which might cripple lysosomal function further—a more surgical approach might target H57 or the salt-bridge network around it. Drugs that stabilize the closed state, or that prevent excessive pH-gradient collapse, could restore balance.
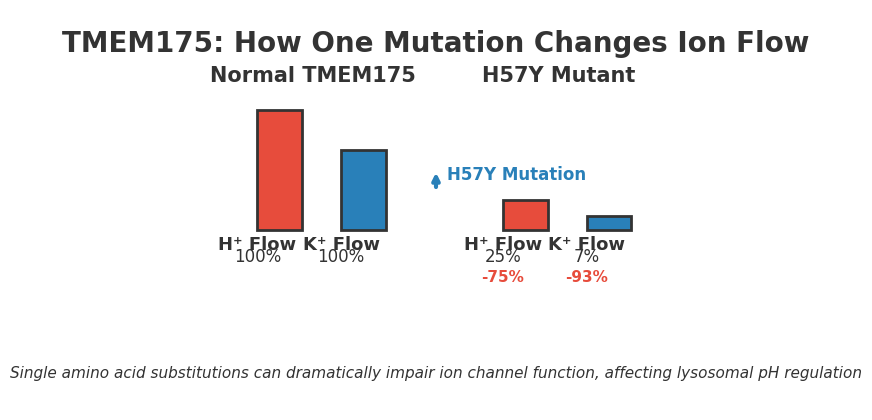
Image file: tmem175-proton-channel.png — Left: TMEM175 channel structure highlighting H57 position on the luminal side and its salt-bridge contacts with D279 and E282. Right: Current traces showing the rapid inward/outward currents and voltage shift when pH jumps from neutral to acidic, demonstrating the channel’s proton-sensing activation.
The Bottom Line
TMEM175 is not a simple K+ channel with a hint of proton permeability. It is a sophisticated acid-sensor that, when triggered, unleashes a torrent of H+ influx potent enough to obliterate the lysosome’s acid gradient in seconds. The H57 residue acts as the molecular switch, and a single mutation can disable the entire system.
For neuroscience, this study cracks open a mechanism long suspected but never clearly shown: how a lysosomal channel linked to Parkinson’s disease actually behaves at the biophysical level. The next challenge is to translate these channel mechanics into cell biology—to understand how H57 variants lead to neurodegeneration in living brains—and ultimately into drugs that can restore balance without breaking the lysosome altogether.
Citation: Schulze T, Sprave T, Groebe C, et al. Proton-selective conductance and gating of the lysosomal cation channel TMEM175. PNAS. 2026;123(3):e2503909123. DOI: 10.1073/pnas.2503909123
Authors’ affiliations: Multiple institutions across Germany and Europe, including the University of Munich (Technical University and Ludwig Maximilian University), the University of Bonn, and others. Correspondence to: christian.grimm@med.uni-muenchen.de or oliver.rauh@h-brs.de