TL;DR: A 2026 study in The Journal of Physiology found that steady-state Purkinje-cell firing patterns had limited ability to predict cerebellar nuclei output in mouse models of cerebellar movement disease.
Key Findings
- 5 disease models: The reanalysis used in vivo recordings from mouse models involving ataxia, dystonia, tremor, and altered cerebellar circuitry.
- Purkinje-to-nuclei test: Researchers tested whether Purkinje-cell spike changes predicted downstream cerebellar nuclei spike changes.
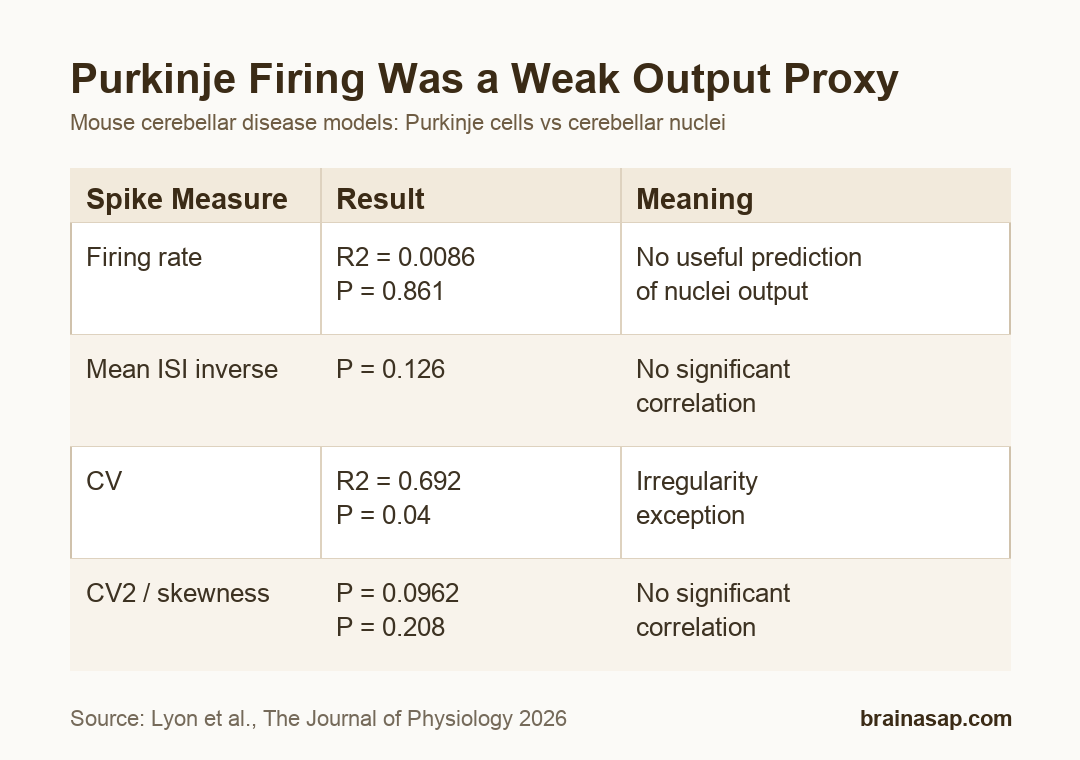
- Firing-rate null: Firing rate showed no meaningful Purkinje-versus-nuclei correlation, with R2 = 0.0086 and P = 0.861.
- CV exception: Coefficient of variation, an irregularity measure, showed a positive correlation with R2 = 0.692 and P = 0.04.
- Therapy caution: The findings warn against assuming that correcting Purkinje-cell firing alone will normalize cerebellar output.
Source: The Journal of Physiology (2026) | Lyon et al.
Purkinje cells are the main inhibitory neurons projecting to the cerebellar nuclei, which provide much of the cerebellum’s output. That wiring makes it tempting to assume that Purkinje firing changes should translate into opposite downstream changes.
This study tested that assumption across disease models instead of accepting it from anatomy alone.
Purkinje Inhibition Did Not Produce a Simple Output Rule
In healthy circuitry, Purkinje cells inhibit cerebellar nuclei neurons. A straightforward model would predict that higher Purkinje firing should lower nuclei firing, while lower Purkinje firing should release nuclei neurons and raise output.
Movement-disorder models complicate that logic because disease can alter firing rate, firing regularity, synaptic wiring, or multiple parts of the circuit at once.
- Purkinje cells: Inhibitory cerebellar cortical output neurons that project to cerebellar nuclei.
- Cerebellar nuclei: Downstream output neurons that influence movement circuits.
- Spike irregularity: Measures such as coefficient of variation describe how variable the timing between spikes becomes.
The authors reanalyzed previously published in vivo electrophysiology datasets rather than recording a new single cohort. Data came from adult mice of mixed sex and background, with each disease model normalized to its own control group.
That design made the study useful for a cross-model question: whether disease-linked Purkinje firing changes provide a reliable population-level readout of nuclei firing changes.
Firing Rate Failed as a Purkinje-to-Nuclei Predictor
The cleanest test was firing rate. If the simple inhibitory model held across disease states, Purkinje and nuclei firing-rate changes should have shown a consistent negative correlation.
They did not. Linear regression for firing rate produced R2 = 0.0086, P = 0.861, and a confidence interval spanning negative and positive slopes.
- Firing rate: No significant correlation between Purkinje-cell and nuclei-cell changes.
- Mean ISI inverse: The interspike-interval rate measure also did not pass the significance threshold.
- Long pauses: The percentage of intervals longer than 25 ms showed no significant relationship.
The null result matters for biomarker thinking. Purkinje firing rate alone cannot be treated as a direct surrogate for cerebellar output across these disease models.
Spike Irregularity Was More Complicated Than Firing Rate
The irregularity measures were not uniformly negative. Coefficient of variation, or CV, showed a significant positive correlation between Purkinje-cell and nuclei-cell changes.
For CV, the table reported R2 = 0.692, P = 0.04, and a slope confidence interval of 0.0477 to 1.2345. The bootstrap analysis found significance in 51% of runs.
- CV: Positive correlation, suggesting some shared irregularity change across the circuit.
- CV2: No significant correlation despite a moderate R2 value.
- Skewness: No significant correlation between Purkinje and nuclei spike-timing changes.
That mixed irregularity profile is important. It suggests that disease-related spike timing can move together in some ways, but not through a single stable rule that every parameter follows.
Disease Models May Change More Than One Cerebellar Circuit Node
The paper included genetic, pharmacological, and circuit-based models. Examples included Car8wdl/wdl, harmaline injection, altered Vglut2 or Vgat signaling, Ank1 deletion, and Sca1-related disease modeling.
Those models can affect Purkinje cells, nuclei cells, upstream inputs, or synaptic properties differently. A downstream nuclei spike pattern may therefore reflect more than the current steady-state firing of Purkinje cells.
- Cell-autonomous effects: Nuclei neurons may change their own excitability in disease.
- Input diversity: Cerebellar nuclei receive more than Purkinje inhibition, including excitatory inputs.
- Timing effects: Firing irregularity may influence output differently from average firing rate.
This helps explain why anatomy alone was not enough. The inhibitory synapse is real, but disease can change the circuit context around that synapse.
Reanalyzed Mouse Data Limit the Treatment Interpretation
The study is strongest as a reanalysis of steady-state spike recordings. It does not test every cerebellar disease mechanism, every behavioral state, or acute therapeutic manipulation in a controlled intervention pipeline.
Some datasets also lacked paired Purkinje and nuclei recordings for every model. In models where Purkinje neurotransmission was minimal, only nuclei-cell recordings were available.
- Data source: Recordings were reanalyzed from earlier mouse studies.
- Model scope: Mouse disease models do not fully represent human ataxia, dystonia, or tremor.
- Therapy boundary: Normalizing Purkinje-cell firing may still help in some contexts, but it should not be assumed to normalize nuclei output.
The practical result is a measurement caution: cerebellar therapies and biomarkers need to verify downstream output directly instead of inferring it from Purkinje firing rate alone.
For experimental design, that means recording nuclei activity remains valuable even when Purkinje recordings are available. The downstream cells can show circuit changes that are not visible from the upstream inhibitory firing rate.
Citation: DOI: 10.1113/JP290000. Lyon et al. Steady-state Purkinje cell activity has limited predictive power for cerebellar output in disease. The Journal of Physiology. 2026;604:3964-3983.
Study Design: Reanalysis of in vivo mouse electrophysiology recordings across cerebellar movement-disorder models.
Sample/Model: Adult mouse models of cerebellar disease with Purkinje-cell and cerebellar nuclei recordings from previously published datasets.
Key Statistic: Firing-rate correlation was absent (R2 = 0.0086, P = 0.861), while CV showed a positive irregularity correlation (R2 = 0.692, P = 0.04).
Caveat: The analysis used mouse disease models and reanalyzed datasets, so it does not prove how human cerebellar symptoms will respond to Purkinje-focused treatments.