TL;DR: A 2026 study in Nature Communications reported that tubulin redirected Tau and alpha-synuclein condensates toward microtubule-building states, while tubulin loss pushed the same protein system toward toxic oligomers in cell models.
Key Findings
- 36-hour shift: Tau and alpha-synuclein condensates formed stable high-molecular-weight oligomers and Tau:alpha-synuclein heterodimers after aging without tubulin.
- 10-micromolar tubulin: Adding tubulin to 20:10 micromolar Tau:alpha-synuclein mixtures transformed round droplets into elongated microtubule-rich tactoids.
- FRET-FLIM lifetimes: Tubulin raised compact-condensate donor lifetimes from about 2.7 to 3.3 ns for Tau and from about 2.8 to 3.2 ns for alpha-synuclein.
- Lower FRET readout: Tubulin-rich condensates shifted Tau and alpha-synuclein from high-FRET compact states toward lower-FRET extended states.
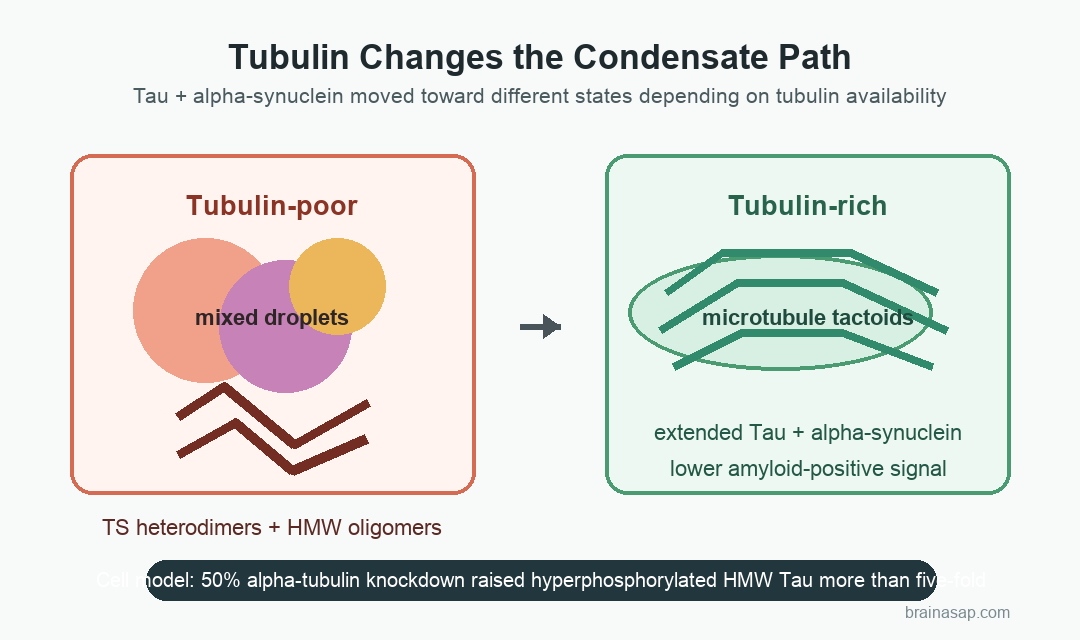
- Cell stress result: Reducing alpha-tubulin by about 50% in N2a cells produced a greater than five-fold rise in hyperphosphorylated high-molecular-weight Tau oligomers.
Source: Nature Communications (2026) | Lucas et al.
Tau and alpha-synuclein are best known through their links to Alzheimer’s disease, Parkinson’s disease, and Lewy body dementia. Both proteins also have ordinary neuronal jobs, including roles near microtubules, the internal tracks that help neurons maintain shape and move cargo.
The difficulty is that the same proteins can gather into biomolecular condensates. Those droplet-like assemblies can support normal cell organization, but they can also mature into aggregate-prone states when the local protein balance changes.
Tau and Alpha-Synuclein Condensates Became Oligomer-Rich Without Tubulin
Researchers first rebuilt the protein system in vitro using purified Tau, alpha-synuclein, and tubulin. The goal was to test whether tubulin simply sits downstream of neurodegeneration or actively changes how Tau and alpha-synuclein behave inside condensates.
The starting point was a Tau-driven liquid-liquid phase separation system. Tau readily formed condensates under the test conditions, while alpha-synuclein alone did not form the same droplets at physiological-like concentrations.
- Tau recruitment: Tau condensates pulled alpha-synuclein into mixed Tau:alpha-synuclein droplets.
- Aging window: Samples were followed over time, including 0-, 16-, and 36-hour incubations for oligomer assays.
- Protein readout: SDS-PAGE, western blots, cross-linking, and LC-MS/MS were used to identify stable oligomer bands.
- Amyloid readout: Dot blots tested whether aged condensates developed amyloid-positive structure.
In tubulin-poor condensates, Tau and alpha-synuclein formed high-molecular-weight oligomers. A Tau:alpha-synuclein heterodimer band, labeled TS in the paper, became more visible with longer incubation and higher alpha-synuclein concentration.
LC-MS/MS supported that identity: the excised TS band contained 22 unique Tau peptides and 3 unique alpha-synuclein peptides, with no other high-confidence protein identification. That made the mixed-protein oligomer assignment stronger than a simple gel-position guess.
Tubulin-Rich Droplets Shifted Toward Microtubule Tactoids
Adding tubulin changed the physical shape of the condensates. Round Tau:alpha-synuclein droplets became elongated tactoids, a spindle-like form linked to microtubule organization.
Time-resolved confocal microscopy tracked mixtures with Tau, alpha-synuclein, and tubulin across concentration ranges from 0 to 20 micromolar. A representative condition used 20 micromolar Tau, 10 micromolar alpha-synuclein, and 10 micromolar tubulin.
Fluorescence recovery after photobleaching, or FRAP, showed that both droplets and tactoids retained liquid-like behavior. Tubulin slowed recovery, consistent with more stable microtubule bundles inside the condensate environment.
- Droplet state: Tau recruited alpha-synuclein into mixed condensates that favored TS heterodimers and amyloid-positive material.
- Tactoid state: Tubulin co-partitioned into condensates and promoted microtubule-like organization.
- Redistribution: Average Tau and alpha-synuclein partitioning decreased as tubulin concentration rose, reflecting a shift in condensate composition and morphology.
In these assays, tubulin marked a healthier cell state and directly redirected the condensate toward a microtubule-associated form.
Western Blots and FRET-FLIM Showed Fewer Oligomers and More Extended Proteins
The biochemical assays supported the microscopy. When tubulin was present, Tau:alpha-synuclein homotypic and heterotypic oligomers were reduced, and the TS heterodimer was barely visible in the condensate pellet fraction.
That effect fits the structural map of Tau. Alpha-synuclein can interact near Tau’s proline-rich and microtubule-binding regions.
Tubulin also binds the microtubule-binding domain. That overlap may let tubulin compete with aggregate-prone Tau:alpha-synuclein contacts.
- Gel assays: Tubulin-containing condensates showed fewer high-molecular-weight Tau and alpha-synuclein species.
- Amyloid assay: Tubulin reduced amyloid-positive signal in the dot blot after long incubation.
- Complexes detected: Soluble Tau:tubulin and alpha-synuclein:tubulin species appeared in western blot data.
- Structural readout: FRET-FLIM tested whether the proteins were compact or extended inside condensates.
FRET-FLIM supplied the most direct structural readout. Without tubulin, Tau donor lifetime shifted from about 3.2 ns in the dilute phase to about 2.7 ns in condensates, indicating compaction.
Alpha-synuclein showed a similar shift from about 3.4 ns to about 2.8 ns.
With tubulin, those lifetimes moved back toward extended states: roughly 3.3 ns for Tau and 3.2 ns for alpha-synuclein. FRET efficiency also dropped, from about 1.0 for compact Tau and 0.9 for compact alpha-synuclein toward about 0.6 and 0.7, respectively.
Tubulin Knockdown Raised Hyperphosphorylated Tau in N2a Cells
The cell experiments tested whether the same direction appeared in a neuronal model. In mouse Neuro-2a cells, siRNA reduced alpha-tubulin expression by about 50%, then researchers measured endogenous Tau, phospho-Tau, and alpha-synuclein oligomerization by western blot.
Tau oligomerization increased after tubulin reduction. The sharper signal came from hyperphosphorylated Tau: high-molecular-weight phospho-Tau oligomers rose by more than five-fold.
Alpha-synuclein oligomers above 100 kDa did not significantly increase in that same assay. The paper interprets that as compatible with delayed alpha-synuclein aggregation or with alpha-synuclein having other cellular functions beyond microtubules.
- Cell model: Mouse N2a cells supplied the tubulin-knockdown assay.
- Replicates: Western blot quantification used 3 biological replicates.
- Stress model: Okadaic acid was used in separate assays to drive Tau hyperphosphorylation and microtubule detachment.
- Neurite readout: Tubulin siRNA and oxidative stress reduced neurite extensions in N2a cells expressing OptoTau and GFP-alpha-synuclein.
Optogenetic Tau experiments added a second angle. In SH-SY5Y cells, blue-light activation of an OptoTau construct promoted microtubule-like structures under okadaic-acid stress, showing that Tau-rich condensates can still support microtubules in the right context.
Microtubule-Stabilizing Ideas Remain Preclinical
The study changes how tubulin can be framed in neurodegeneration models. Instead of treating microtubule loss only as damage that happens after aggregation, the data suggest that tubulin availability can shape whether Tau and alpha-synuclein condensates become functional or oligomer-prone.
That does not make tubulin restoration a ready therapy. Microtubule-stabilizing drugs can have broad effects, and some agents may displace Tau from microtubules in ways that could worsen aggregation risk.
- Model boundary: The work used purified proteins, mouse N2a cells, human SH-SY5Y cells, and engineered optogenetic constructs.
- Disease boundary: Alzheimer’s disease and Parkinson’s disease relevance comes from shared Tau, alpha-synuclein, and microtubule biology, not from patient intervention data.
- Therapy boundary: The data support testing selective ways to preserve tubulin-rich condensate states, not indiscriminate condensate blockade.
The strongest supported claim is mechanistic: tubulin-rich environments pushed Tau and alpha-synuclein toward extended, microtubule-associated states, while tubulin loss favored Tau oligomerization, phospho-Tau accumulation, and neurite loss in cell models.
Citation: DOI: 10.1038/s41467-026-69618-3. Lucas et al. Tubulin transforms Tau and alpha-synuclein condensates from pathological to physiological. Nature Communications. 2026.
Study Design: Mechanistic preclinical study combining purified-protein phase-separation assays, microscopy, western blotting, amyloid dot blots, FRET-FLIM, and neuronal cell models.
Sample/Model: Purified Tau, alpha-synuclein, and tubulin systems; mouse N2a cells; human SH-SY5Y neuroblastoma cells; OptoTau and GFP-alpha-synuclein constructs.
Key Statistic: About 50% alpha-tubulin knockdown in N2a cells produced a greater than five-fold increase in hyperphosphorylated high-molecular-weight Tau oligomers.
Caveat: The findings are mechanistic and preclinical; they do not show that a tubulin-targeted treatment prevents or slows human neurodegenerative disease.





